Topp-ti problemer med evolusjon: 2. Homologi
Oversatt herfra.
Redaktørens notat: Vi er glade for å presentere en serie av biolog Jonathan Wells om de viktigste vitenskapelige problemene med evolusjon. Dette er den andre oppføringen i serien, utdrag fra den nye boken The Comprehensive Guide to Science and Faith: Exploring the Ultimate Questions About Life and the Cosmos -lenke . Finn hele serien, så langt her.
Klassisk betydde av homologi er likhet i struktur og posisjon: for eksempel i beinene i den menneskelige hånden og vingen til en flaggermus. Darwin anså homologier for å være bevis på felles aner. Han skrev i On the Origin of Species:
-Hva kan være mer rart enn at hånden til en mann, formet for å gripe, den til en føflekk for å grave, beinet til hesten, finnen til nisen og vingen til flaggermusen, alle skal være bygget på samme mønster, og bør inkludere de samme beinene, i samme relative posisjoner?(1) {Unntatt for dem som regner med samme designer for dette -oversetters tilføyelse.}
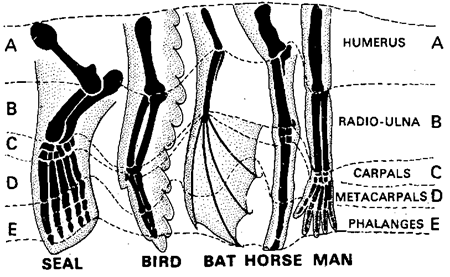 Darwin betraktet dette som uforklarlig hvis alle arter ble skapt separat: "På det vanlige synet på den uavhengige skapelsen av hvert vesen, kan vi bare si at det er slik; -- at det har behaget Skaperen så mye å konstruere hvert dyr og plante."(2) (I den fjerde utgaven av On the Origin of Species la Darwin til "men dette er ikke en vitenskapelig forklaring."(3) I stedet, hevdet han, var homologier forklarlig med hans hypotese om avstamning med modifikasjon:
Darwin betraktet dette som uforklarlig hvis alle arter ble skapt separat: "På det vanlige synet på den uavhengige skapelsen av hvert vesen, kan vi bare si at det er slik; -- at det har behaget Skaperen så mye å konstruere hvert dyr og plante."(2) (I den fjerde utgaven av On the Origin of Species la Darwin til "men dette er ikke en vitenskapelig forklaring."(3) I stedet, hevdet han, var homologier forklarlig med hans hypotese om avstamning med modifikasjon:
-Hvis vi antar at den eldgamle stamfaderen, arketypen som den kan kalles, av alle pattedyr, hadde sine lemmer konstruert etter det eksisterende generelle mønsteret, uansett formål de tjente, kan vi med en gang oppfatte den enkle betydningen av den homologe konstruksjonen av lemmer gjennom hele klassen.(4)
Kameraøyne, pigger og mer
Likevel har dyr og planter mange egenskaper som er like i struktur og posisjon, men er tydeligvis ikke avledet fra en felles stamfar med disse egenskapene. Kameraøyet til et virveldyr og kameraøyet til en blekksprut eller blekksprut er bemerkelsesverdig like, men ingen tror de ble arvet fra en felles stamfar som hadde et kameraøye. Piggene til australske piggsvin og nordamerikanske piggsvin er bemerkelsesverdig like, men echidnas-piggsvin føder ved å legge egg, mens porcupines-piggsvin føder levende babyer, etter å ha gitt dem næring i livmoren, lik mennesker. Denne grunnleggende forskjellen betyr at de to typene pggsvin hadde svært forskjellig opprinnelse, og de arvet ikke pigger fra en pigget felles stamfar. Hudfoldene mellom forbenene og bakbenene til australske flygende phalangers og nordamerikanske flygende ekorn er svært like. Likevel føder førstnevnte fostre som kryper inn i en pose for å fullføre utviklingen, lik kenguruer, mens de sistnevnte gir fostrene sine i livmoren, som 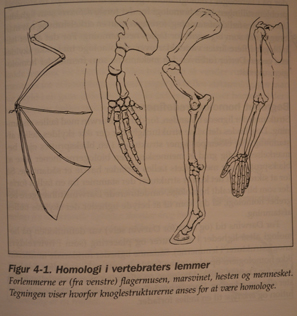 mennesker. Igjen hadde de svært forskjellig opphav.(5)
mennesker. Igjen hadde de svært forskjellig opphav.(5)
Eksempler inkluderer også gropene av klebrig nektar i kjøttetende planter, som tilsynelatende har sin opprinnelse hver for seg, seks forskjellige ganger.(6) Planter av slekten euphorbia i Afrika har tykke, kjøttfulle stengler for å lagre vann og stikkende pigger i stedet for blader, som planter av kaktusfamilien i Amerika, men de oppsto hver for seg under svært forskjellige forhold.(7)
Bortsett fra når det ikke er det
Så likhet i struktur og posisjon er bevis for felles aner, bortsett fra når det ikke er det. Moderne biologer kaller likhet ikke på grunn av felles aner, for konvergens, og de har omdefinert homologi til å bety likhet på grunn av felles aner. Berkeleys evolusjonsbiolog David Wake skrev i 1999, "Felles aner er alt som er til homologi."(8) Men ifølge biologifilosofen Ronald Brady, "Ved å gjøre vår forklaring [felles aner] til definisjonen av tilstanden som skal forklares [homologi] ], uttrykker vi ikke vitenskapelige hypoteser, men tro."(9)
 Resonnering i en sirkel
Resonnering i en sirkel
Videre, når homologi er definert i form av felles aner, kan den ikke logisk brukes som bevis for felles aner. å gjøre det vil være å resonnere i en sirkel: Hvordan vet vi at trekk A og trekk B stammer fra en felles stamfar? Fordi de er homologe. Hvordan vet vi at A og B er homologe? Fordi de stammer fra en felles stamfar.(10)
Et annet problem med å bruke homologi som bevis for felles aner er at eksempler på konvergens er utbredt. Cambridge paleobiolog Simon Conway Morris skrev i 2003 at "konvergens er allestedsnærværende." Han konkluderte med: "Ikke bare er universet merkelig egnet til formålet, men det er også livets evne til å navigere til dets løsninger."(11) Så likheter i struktur og posisjon gir ikke klare bevis for det felles forfedreaspektet ved evolusjon.
Referanser:
1. Charles Darwin, Origin of Species, 1st ed. (1859), 434, http://darwin-online.org.uk/content/frameset?pageseq=453&itemID=F373&viewtype=side (accessed August 23, 2020).
2. Darwin, Origin of Species, 1st ed., 435.
3. Charles Darwin, Origin of Species, 4th ed. (1866), 513, http://darwin-online.org.uk/content/frameset?pageseq=545&itemID=F385&viewtype=side (accessed August 23, 2020).
4. Darwin, Origin of Species, 1st ed., 435.
5. Jonathan Wells, Zombie Science: More Icons of Evolution (Seattle, WA: Discovery Institute Press, 2017), 44-47.
6. Thomas J. Givnish, "New evidence on the origin of carnivorous plants," Proceedings of the National Academy of Sciences USA 112 (2015), 10-11.
7. Leonardo O. Alvarado-Cárdenas, Enrique Martínez-Meyer, Teresa P. Feria, Luis E. Eguiarte, Héctor M. Hernández, Guy Midgley, and Mark E. Olson, "To converge or not to converge in environmental space: Testing for similar environments between analogous succulent plants of North America and Africa," Annals of Botany 111 (2013), 1125-1138.
8. David B. Wake, "Homoplasy, homology and the problem of ‘sameness’ in biology," Novartis Symposium 222--Homology, eds. G.K. Bock and G. Cardew (Chichester, UK: John Wiley & Sons, 1999), 45.
9.Ronald H. Brady, "On the independence of systematics," Cladistics 1 (1985), 113-126.
10. Wells, Zombie Science, 42.
11. Simon Conway Morris, Life’s Solution: Inevitable Humans in a Lonely Universe (Cambridge, NY: Cambridge University Press, 2003), 283, 327.
Neste artikkel: Problemet med fossiler -under her:
---
Topp-ti problemer med evolusjon: 3. Fossiler
Oversatt herfra.
Redaktørens notat: Vi er glade for å presentere en serie av biolog Jonathan Wells om de viktigste vitenskapelige problemene med evolusjon. Dette er den tredje oppføringen i serien, utdrag fra den nye boken The Comprehensive Guide to Science and Faith: Exploring the Ultimate Questions About Life and the Cosmos -lenke . Finn hele serien, så langt her.
 Et fossil er "en rest, et inntrykk eller et spor av en organisme fra tidligere geologiske tidsaldre".(1) Studiet av fossiler (kalt paleontologi) startet lenge før Darwin. De gir vårt beste glimt av livets historie før nåtiden. Forutsatt at fossiler i ett lag av stein er yngre enn fossiler i lag under dem, hadde før-darwinstiske paleontologer allerede gruppert dem i henhold til deres relative alder. Resultatet er kjent som fossilregisteret.
Et fossil er "en rest, et inntrykk eller et spor av en organisme fra tidligere geologiske tidsaldre".(1) Studiet av fossiler (kalt paleontologi) startet lenge før Darwin. De gir vårt beste glimt av livets historie før nåtiden. Forutsatt at fossiler i ett lag av stein er yngre enn fossiler i lag under dem, hadde før-darwinstiske paleontologer allerede gruppert dem i henhold til deres relative alder. Resultatet er kjent som fossilregisteret.
Darwin skrev dette om fossilregisteret i On the Origin of Species:
Ved teorien om naturlig utvalg har alle levende arter blitt forbundet med foreldreartene til hver slekt, ved forskjeller som ikke er større enn vi ser mellom variantene av samme art i dag; og disse foreldreartene, som nå generelt er utdødd, har i sin tur, på samme måte vært forbundet med eldre arter; og så videre bakover, alltid konvergerende til den felles stamfaren til hver stor klasse. Slik at antallet mellom- og overgangsledd, mellom alle levende og utdødde arter, må ha vært ufattelig stort.(2)
Men det 'ufattelig store' antallet overgangskoblinger postulert av Darwin, har aldri blitt funnet. Faktisk er et av de mest fremtredende trekkene ved fossilregistreringen den kambriske eksplosjonen, der hovedgruppene av dyr (kalt phyla) dukket opp rundt samme geologiske tid i en periode kalt kambrium, fullt utformet og uten fossile bevis på at de divergerte fra en felles stamfar.
Relaterte Artikler
-
- Jonathan Wells, 14.Februar, 2022
-
-
- Jonathan Wells, 10. February, 2022
-
- David Klinghoffer, 8. December 2021
Et alvorlig problem for teorien
Darwin visste om disse bevisene i 1859, og han erkjente at det var et alvorlig problem som "virkelig kan være et gyldig argument" mot hans teori.(3) Han håpet at fremtidige fossilfunn ville bidra til å fylle ut mange av tomrommene, men mer enn 150 år med ekstra fossilinnsamling har bare gjort problemet verre. I 1991 konkluderte et team av paleontologer at den kambriske eksplosjonen "var enda mer brå og omfattende enn tidligere antatt." (4)
Det plutselige som ble observert i den kambriske eksplosjonen, kan også sees i mindre skalaer gjennom hele fossilregistret. Arter har en tendens til å dukke opp brått i fossilregisteret og deretter vedvare uendret i en periode (et fenomen som kalles stasis) før de forsvinner. I 1972 kalte paleontologene Niles Eldredge og Stephen Jay Gould dette mønsteret for punktert likevekt.(5) I følge Gould visste "hver paleontolog" at det er det dominerende mønsteret i fossilregisteret.(6) Med andre ord, det "alltid ufattelig store" antallet av overgangskoblinger postulert av Darwin, ikke bare mangler i den kambriske eksplosjonen, men gjennom hele fossilhistorien.
 To menneskelige skjeletter
To menneskelige skjeletter
Selv om vi hadde en god fossilhistorie, ville vi fortsatt trenge vår fantasi for å produsere fortellinger om relasjoner mellom forfedre og etterkommere. Her er grunnen: Hvis du fant to menneskeskjeletter begravd i et felt, hvordan kunne du vite om det ene stammet fra det andre? Uten identifiseringsmerker og skriftlige dokumenter, eller kanskje i noen tilfeller DNA, ville det være umulig å vite. Likevel vil du ha å gjøre med to skjeletter fra samme nyere, levende art. Med to forskjellige, eldgamle, utdødde arter - ofte langt fra hverandre i tid og rom - ville det ikke vært mulig å demonstrere et forfedre-etterkommer forhold.
For flere tiår siden skrev paleontolog Gareth Nelson, "Ideen om at man kan gå til fossilregistrene og forvente å empirisk gjenopprette en forfedre-etterkommersekvens, det være seg arter, slekter, familier eller hva som helst, har vært, og fortsetter å være, en skadelig illusjon."(7) I 1999 skrev evolusjonsbiolog Henry Gee at "det er faktisk umulig å koble fossiler til kjeder av årsak og virkning på noen gyldig måte". Han konkluderte: "å ta en linje med fossiler og hevde at de representerer en avstamning, er ikke en vitenskapelig hypotese som kan testes, men en påstand som bærer samme gyldighet som en godnatt-historie - morsomt, kanskje til og med lærerikt, men ikke vitenskapelig."(8)
Neste artikkel. Molekylær fylogeni -rett under her:
Referanser:
-
Merriam-Webster’s definition of "fossil," https://www.merriam-webster.com/dictionary/fossil (accessed August 23, 2020).
-
-
-
James W. Valentine, Stanley M. Awramik, Philip W. Signor, and Peter M. Sadler, "The biological explosion at the Precambrian-Cambrian boundary," Evolutionary Biology 25 (1991), 279-356.
-
Niles Eldredge and Stephen Jay Gould, "Punctuated equilibria: An alternative to phyletic gradualism," Models in Paleobiology, ed. Thomas J. M. Schopf (San Francisco, CA: Freeman Cooper, 1972), 82-115.
-
Stephen Jay Gould, The Structure of Evolutionary Theory (Cambridge, MA: Harvard University Press, 2002), 759.
-
Gareth Nelson, "Presentation to the American Museum of Natural History" (1969), in David M. Williams and Malte C. Ebach, "The reform of palaeontology and the rise of biogeography," Journal of Biogeography 31 (2004), 685-712.
-
Henry Gee, In Search of Deep Time: Beyond the Fossil Record to a New History of Life (New York: The Free Press, 1999), 113, 116-117.
---
Topp-ti problemer med evolusjon: 4. Molekylær fylogeni
Oversatt herfra.
 Redaktørens notat: Vi er glade for å presentere en serie av biolog Jonathan Wells om de viktigste vitenskapelige problemene med evolusjon. Dette er den fjerde oppføringen i serien, utdrag fra den nye boken The Comprehensive Guide to Science and Faith: Exploring the Ultimate Questions About Life and the Cosmos -lenke . Finn hele serien, så langt her.
Redaktørens notat: Vi er glade for å presentere en serie av biolog Jonathan Wells om de viktigste vitenskapelige problemene med evolusjon. Dette er den fjerde oppføringen i serien, utdrag fra den nye boken The Comprehensive Guide to Science and Faith: Exploring the Ultimate Questions About Life and the Cosmos -lenke . Finn hele serien, så langt her.
Ordet fylogeni refererer til den evolusjonære historien til en organisme.(1) Ordet ble laget av den tyske darwinistiske biologen Ernst Haeckel flere år etter utgivelsen av On the Origin of Species. Evolusjonsbiologer har foreslått fylogenier basert på homologier i fossiler, men som vi har sett er det problemer med både fossiler og homologi. Med fremveksten av moderne molekylærbiologi har evolusjonsbiologer i økende grad forsøkt å basere fylogenier på molekyler som proteiner og DNA.
Proteiner består av sekvenser av underenheter kalt aminosyrer, og DNA består av underenheter kalt nukleotider. Ulike arter kan inneholde lignende proteiner eller DNA-molekyler som viser små forskjeller i sekvensene til underenhetene deres. Hvis tre forskjellige arter inneholder et lignende DNA-molekyl, og sekvensen i art A er mer lik sekvensen i art B enn i art C, kan en evolusjonsbiolog slutte at A er nærmere beslektet med B enn den er til C.
Definere 'relatert'
Men betydningen av relatert er tvetydig. På en måte kan det referere til slektsforskning, som i "Charles Darwin var nærmere beslektet med Erasmus Darwin (hans bestefar) enn noen av dem var til Geronimo." I en annen forstand kan det referere til likhet, som i "jern er nærmere beslektet med aluminium enn begge er til en påskelilje."(2) Fylogenetiske slutninger antar at molekylær slektskap (den andre betydningen) er ekvivalent med genealogisk slektskap (den første betydnigen). Dette premisset er basert på antakelsen om felles avstamning.
Molekylære sammenligninger kompliseres av problemet med justering. DNA-sekvenser i levende ting inneholder vanligvis gjentatte og/eller slettede segmenter, så det er ofte uklart hvor de skal settes opp. Hvis to sekvenser kan justeres på mer enn én måte, vil enhver sammenligning avhenge sterkt av hvilken justering forskeren velger. Og når mange sekvenser sammenlignes, slik de er i molekylære fylogenier, blir problemet mye verre.(3)
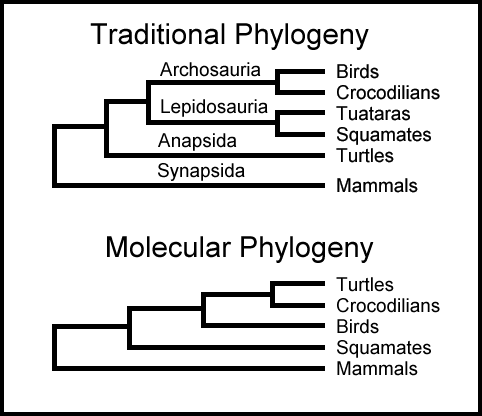 Evolusjonens Livets Tre
Evolusjonens Livets Tre
Darwin mente at levende tings historie kunne representeres som et "stort livstre", med felles forfedre som stammen og moderne organismer som tuppen av grenene.(4) Hvis livets historie er trelignende, ville man forvente at data fra molekylær fylogeni til slutt ville konvergere på et enkelt tre, og at etter hvert som flere data ble funnet, ville tilpasningen bli bedre. Likevel har molekylær fylogenetikk helt fra begynnelsen vært plaget med uoverensstemmelser mellom trær basert på forskjellige sekvenser og forskjellige justeringer.
Og problemet har bare blitt verre ettersom mer data har samlet seg. I 2005 konkluderte tre biologer som sammenlignet 50 DNA-sekvenser fra 17 dyregrupper at "ulike fylogenetiske analyser kan nå motstridende slutninger med [tilsynelatende] absolutt støtte."(5) I 2012 rapporterte fire evolusjonsbiologer "inkongruens mellom fylogenier avledet fra ... forskjellige undergrupper molekylære sekvenser har blitt gjennomgripende."(6)
Så ideen om felles aner forblir en antagelse. Det følger ikke av homologi, bortsett fra ved sirkulært resonnement. Fossilregisteret er fortsatt (som Darwin erkjente), et alvorlig problem. Og felles aner kommer ikke frem fra de inkonsekvente funnene fra molekylær fylogenetikk.
Neste artikkel. Naturlig seleksjon -rett under her:
Referanser
-
-
Wells, Zombie Science, 32-33.
-
James A. Lake, "The order of sequence alignment can bias the selection of tree topology," Molecular Biology and Evolution 8 (1991), 378-385; Wells, Zombie Science, 35-36.
-
-
Antonis Rokas, Dirk Krüger, and Sean B. Carroll, "Animal evolution and the molecular signature of radiations compressed in time," Science 310 (2005), 1933-1938.
-
Liliana Dávalos, Andrea Cirranello, Jonathan Geisler, and Nancy Simmons, "Understanding phylogenetic incongruence: Lessons from phyllostomid bats," Biological Reviews of the Cambridge Philosophical Society 87 (2012), 991-1024.
---
Topp-ti problemer med evolusjon: 5. Naturlig seleksjon
Redaktørens notat: Vi er glade for å presentere en serie av biolog Jonathan Wells om de viktigste vitenskapelige problemene med evolusjon. Dette er den femte oppføringen i serien, utdrag fra den nye boken The Comprehensive Guide to Science and Faith: Exploring the Ultimate Questions About Life and the Cosmos -lenke . Finn hele serien, så langt her.
I introduksjonen til On the Origin of Species skrev Darwin: "Jeg er fullstendig overbevist om at arter ikke er uforanderlige." Han fortsatte: "Videre er jeg overbevist om at naturlig seleksjon har vært den viktigste, men ikke eksklusive måten å modifisere." (1)
Men Darwin hadde ingen bevis for naturlig utvalg. I 'On the Origin of Species' var det beste han kunne tilby 'en eller to imaginære illustrasjoner'.(2) Så i stedet for direkte bevis for naturlig utvalg, baserte Darwin (som selv oppdrettet duer) sin argumentasjon på oppdrett, eller det som ofte kalles kunstig utvalg. Han bemerket at "avlen av husdyr ble nøye ivaretatt i oldtiden", og at "dens betydning består i den store effekten av akkumulering i én retning, i løpet av påfølgende generasjoner, av forskjeller som absolutt ikke er merkbar for et utrent øye". (3)
Artenes opprinnelse
Likevel i alle årene med innenlands avl rapporterte ingen om opprinnelsen til en ny art, langt mindre et nytt organ eller kroppsplan. På 1930-tallet brukte den nydarwinistiske biologen Theodosius Dobzhansky ordet mikroevolusjon for å referere til endringer i eksisterende arter (som de som ble observert av innenlandske oppdrettere), og ordet makroevolusjon for å referere til opprinnelsen til nye arter, organer og kroppsplaner. Han skrev:
-Det er ingen vei mot en forståelse av mekanismene for makroevolusjonære endringer, som krever tid på en geologisk skala, annet enn gjennom en full forståelse av de mikroevolusjonære prosessene som kan observeres i løpet av en menneskelig levetid og ofte kontrollert av menneskets vilje. Av denne grunn er vi tvunget på det nåværende kunnskapsnivået motvillig til å sette et tegn på likhet mellom mekanismene for makro- og mikroevolusjon, og fortsetter på denne antagelsen, for å skyve undersøkelsene våre så langt fremover som denne arbeidshypotesen tillater.(4)
Bevis for naturlig utvalg?
 Men en 'arbeidshypotese' er ikke bevis. Det var ikke før på 1950-tallet at den britiske naturforskeren Bernard Kettlewell oppdaget det som så ut til å være det første beviset for naturlig utvalg. Nattsvermer i Storbritannia finnes hovedsakelig i to varianter: mørk ('melanisk') og lys. Før den industrielle revolusjonen på 1800-tallet var melaniske former sjeldne eller fraværende, men når røyk fra industribyer mørknet nærliggende trestammer, ble den melaniske formen mye mer vanlig. Dette fenomenet, kalt industriell melanisme, ble tilskrevet melaniske møll som var bedre kamuflert enn lette møll og dermed mindre synlig for rovfugler: med andre ord til naturlig utvalg.
Men en 'arbeidshypotese' er ikke bevis. Det var ikke før på 1950-tallet at den britiske naturforskeren Bernard Kettlewell oppdaget det som så ut til å være det første beviset for naturlig utvalg. Nattsvermer i Storbritannia finnes hovedsakelig i to varianter: mørk ('melanisk') og lys. Før den industrielle revolusjonen på 1800-tallet var melaniske former sjeldne eller fraværende, men når røyk fra industribyer mørknet nærliggende trestammer, ble den melaniske formen mye mer vanlig. Dette fenomenet, kalt industriell melanisme, ble tilskrevet melaniske møll som var bedre kamuflert enn lette møll og dermed mindre synlig for rovfugler: med andre ord til naturlig utvalg.
Kettlewell fanget noen av hver variant og merket dem med en liten flekk med maling. Så slapp han dem på mørke eller lyse trestammer. Da han gjenfanget noen dagen etter, fant han ut at en betydelig større andel bedre kamuflerte møll overlevde. Kettlewell kalte dette dette "Darwins manglende bevis."(5) Historien, vanligvis illustrert med bilder av lyse og mørke fargede møll på lyse og mørke trestammer, ble omtalt i flere tiår i mange biologilærebøker som overbevisende bevis for evolusjon .(6)
Nattsvermerens vaner
På 1980-tallet var det imidlertid blitt klart at peprede møll vanligvis ikke hviler på trestammer i naturen. De flyr om natten og hviler om dagen i øvre grener der de ikke kan sees. Ved å slippe ut møll på trestammer på dagtid, klarte ikke Kettlewells eksperiment å simulere naturlige forhold. Det viste seg at de fleste lærebokfotografier hadde blitt iscenesatt ved å feste døde møll på trestammer eller ved å plassere levende møll i unaturlige posisjoner og fotografere dem før de fløy bort.(7)
Bedre bevis for naturlig utvalg kom fra finker på Galápagosøyene på 1970-tallet. øyene var hjemsted for det biologer oppførte som 13 forskjellige arter av finker, og biologene Peter og Rosemary Grant og deres kolleger studerte en av disse på en enkelt øy. Grants  og deres kolleger førte detaljerte registreringer av hver finkeart sin anatomi, inkludert lengden og dybden på nebbet. Da en alvorlig tørke i 1977 tok livet av mange av øyenes planter, døde omtrent 85 prosent av fuglene. Grants og deres kolleger bemerket at de overlevende hadde nebb som i gjennomsnitt var 5 prosent større enn bestandsgjennomsnittet før tørken, antagelig fordi de overlevende fuglene var bedre i stand til å knekke de seige frøene etter tørken. Skiftet skyldtes med andre ord naturlig utvalg. Grants estimerte at hvis en lignende tørke oppstod hvert tiende år, ville fuglenebbet fortsette å bli større, til de ville kvalifisere som en ny art om 200 år.(8)
og deres kolleger førte detaljerte registreringer av hver finkeart sin anatomi, inkludert lengden og dybden på nebbet. Da en alvorlig tørke i 1977 tok livet av mange av øyenes planter, døde omtrent 85 prosent av fuglene. Grants og deres kolleger bemerket at de overlevende hadde nebb som i gjennomsnitt var 5 prosent større enn bestandsgjennomsnittet før tørken, antagelig fordi de overlevende fuglene var bedre i stand til å knekke de seige frøene etter tørken. Skiftet skyldtes med andre ord naturlig utvalg. Grants estimerte at hvis en lignende tørke oppstod hvert tiende år, ville fuglenebbet fortsette å bli større, til de ville kvalifisere som en ny art om 200 år.(8)
De sterkestes ankomst
Da tørken tok slutt og regnet kom tilbake, var det imidlertid rikelig med mat, og den gjennomsnittlige nebbstørrelsen ble normal igjen. Ingen netto evolusjon hadde funnet sted.(9) Likevel fant "Darwins finker" veien inn i de fleste biologilærebøker som bevis for evolusjon ved naturlig utvalg.(10)
Så det er bevis for naturlig utvalg, men i likhet med husdyravl har det aldri blitt observert å produsere noe mer enn mikroevolusjon. Som den nederlandske botanikeren Hugo de Vries skrev i 1904, "Naturlig utvalg kan forklare overlevelsen til de sterkeste, men det kan ikke forklare ankomsten til de sterkeste."(11)
For ankomsten av de sterkeste er de fleste moderne evolusjonsbiologer avhengige av mutasjoner.
Referanser:
1. Charles Darwin, Origin of Species, 1st ed., 6, http://darwin-online.org.uk/content/frameset?pageseq=21&itemID=F373&viewtype=side (accessed August 23, 2020).
2. Darwin, Origin of Species, 1st ed., 90, http://darwin-online.org.uk/content/frameset?pageseq=105&itemID=F373&viewtype=side (accessed August 23, 2020).
3. Darwin, Origin of Species, 1st ed., 32-34, http://darwin-online.org.uk/content/frameset?pageseq=47&itemID=F373&viewtype=side (accessed August 23, 2020).
4. Theodosius Dobzhansky, Genetics and the Origin of Species (New York: Columbia University Press, 1937), 12.
5. H.B.D. Kettlewell, "Darwin’s missing evidence," Scientific American 200 (1959), 48-53.
6. Jonathan Wells, "Second Thoughts About Peppered Moths: This classical story of evolution by natural selection needs revising," The Scientist 13 (May 24, 1999), https://www.discovery.org/a/590/ (accessed August 23, 2020); Jonathan Wells, Icons of Evolution (Washington, DC: Regnery, 2000), 137-157.
7. Judith Hooper, Of Moths and Men: Intrigue, Tragedy and the Peppered Moth (London, UK: Fourth Estate, 2002); Wells, Zombie Science, 63-66.
8, Peter T. Boag and Peter R. Grant, "Intense natural selection in a population of Darwin’s finches (Geospizinae) in the Galápagos," Science 214 (1981), 82-85.
9. H. Lisle Gibbs and Peter R. Grant, "Oscillating selection on Darwin’s finches," Nature 327 (1987), 511-513.
10. Wells, Icons of Evolution, 159-175.
11.Hugo de Vries, Species and Varieties, Their Origin by Mutation, 2d ed. (Chicago, IL: Open Court Press, 1906), 825-826, https://www.gutenberg.org/files/7234/7234-h/7234-h.htm (accessed August 23, 2020).
Oversettelse og bilder ved Asbjørn E. Lund Top Scientific Problems with Evolution: Molecular Phylogeny
Top Scientific Problems with Evolution: Molecular Phylogeny Oversatt
Oversatt  Neste gang:
Neste gang: